
一、研究背景:植物RNA干扰的"双刃剑"
在微观世界中,植物与病毒的战争已持续数亿年。当病毒入侵植物细胞,释放其RNA基因组时,植物并非被动挨打——它们进化出一套精密的"RNA防御系统",其中小干扰RNA(siRNA)扮演着"分子导弹"的角色,精准识别并摧毁病毒RNA。
在拟南芥等模式植物中,DICER-LIKE 4(DCL4)与辅助蛋白DRB4组成的复合物,是生产21核苷酸(21-nt)siRNA的核心"工厂"。这些siRNA如同定制的"分子钥匙",能特异性识别病毒RNA序列,引导降解机器精准摧毁入侵者。过去二十年,科学家们知道DCL4-DRB4对长RNA底物有独特偏好,却始终未能揭开其分子工作机制的面纱。
2026年2月,南方科技大学杜嘉木团队在《Nature Plants》发表里程碑式研究,首次解析了DCL4-RNA复合物在切割活性构象下,以及DCL4-DRB4-RNA复合物在预切割构象下的高分辨率结构,揭示了植物如何精准"裁剪"21-nt siRNA的分子密码。这项由中国科学家主导的研究,不仅填补了植物RNA干扰领域的关键知识空白,更开启了基于结构设计的作物保护新策略。
二、突破性发现
1. 精准的"分子尺":21-nt siRNA如何被精确测量?
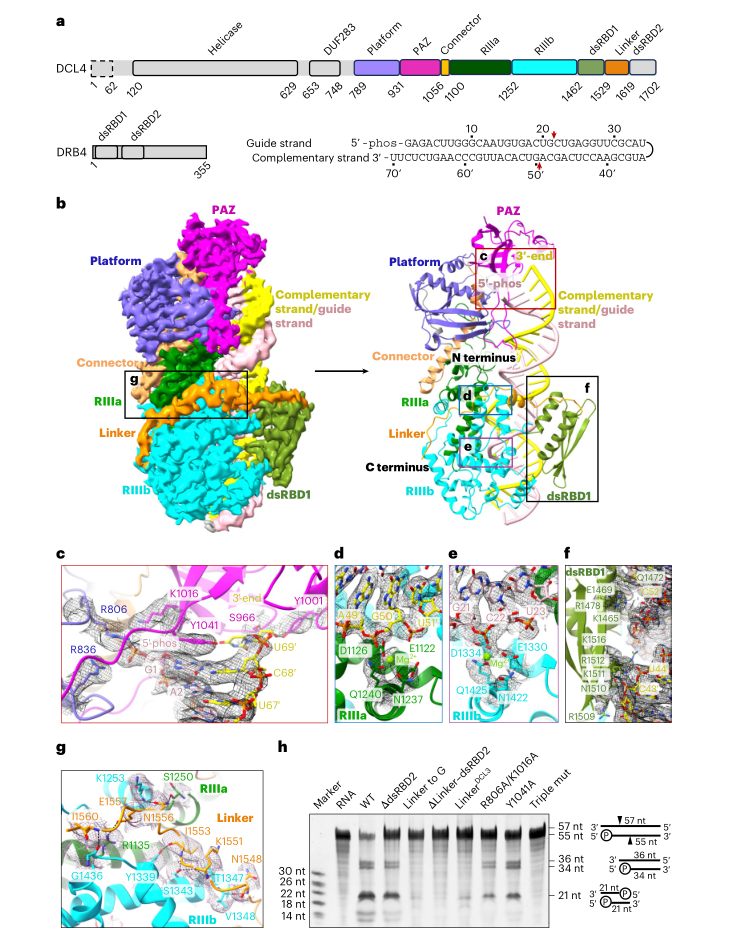
研究团队首先捕获了DCL4-RNA复合物在切割活性构象下的3.15Å分辨率结构。令人惊叹的是,DCL4像一位精准的裁缝,通过其PAZ结构域与RNase III结构域之间的固定距离,沿RNA底物精确测量21个核苷酸的长度。
结构显示:DCL4 PAZ结构域的Tyr1041残基与引导链RNA 5'-G1碱基形成堆叠作用,成为测量的起始点;而RNase IIIb结构域的活性位点则精准切割引导链上G21与C22之间的磷酸二酯键,确保生成严格21-nt的siRNA。这种"起始-终止"的分子测量机制,解释了为何DCL4产生的siRNA长度高度一致,而这一特性对于siRNA有效装载到AGO蛋白并发挥功能至关重要。
2. 独特的"长RNA偏好":植物抗病毒的关键适应
与哺乳动物Dicer不同,DCL4对RNA底物表现出明显的长度偏好——它高效切割50-500 bp的长双链RNA(dsRNA),而非短片段。这项研究首次从结构上揭示了这一现象的分子基础:
LinkerRBDs区域的关键作用:DCL4特有的一个长环状区域(LinkerRBDs)包裹在RNase III结构域周围,将dsRBD2结构域(双链RNA结合结构域2)定位到RNA底物的远端位置(距5'端约50 bp处)。这意味着DCL4需要至少50 bp的RNA才能实现完整的底物结合,从结构上解释了其长RNA偏好性。
DRB4的协同定位:在DCL4-DRB4-RNA预切割构象结构中,DRB4通过其C端结构域(DRB4CTD)与DCL4的dsRBD2相互作用,进一步将DRB4的两个N端dsRBD结构域导向RNA更远端的位置。这种"手拉手"的协同作用,极大扩展了复合物对长RNA的捕获能力。
实验验证:体外切割实验证实,当LinkerRBDs被替换为甘氨酸链或删除时,DCL4的切割活性显著降低;而完整DCL4-DRB4复合物对107/109-nt RNA的切割效率远高于30/32-nt RNA,印证了结构观察。
3. 底物识别的"宽容哲学":适应多样病毒入侵
研究揭示了DCL4在底物识别上的独特策略——对关键末端特征的低选择性:
这种"宽容"的底物识别策略,使DCL4-DRB4复合物能应对多样化的病毒RNA底物,也为连续切割产生相位化siRNA(phasiRNA)提供了基础——在首次切割产生2-nt 3'突出后,该突出恰好成为下一轮切割的理想底物特征,实现连续"流水线"式siRNA生产。
4. 从捕获到切割:RNA加工的动态循环
通过比较DCL4在预切割与切割活性两种构象下的结构,研究团队提出了DCL4-DRB4加工长RNA的动态模型:
捕获阶段:DRB4协助DCL4捕获长dsRNA,DCL4的解旋酶结构域像"马达"一样沿RNA移动
定位阶段:当RNA末端到达PAZ结构域时,移动暂停,RNA在C29位置弯曲
切割阶段:RNA进一步弯曲至C11位置,接近RNase III活性中心,进行精确切割
循环阶段:21-nt siRNA释放后,解旋酶结构域将剩余RNA"泵"向下一个切割位点
这一"抓取-弯曲-切割-移动"的循环机制,完美解释了DCL4如何在长RNA上连续产生等间距的21-nt siRNA,形成phasiRNA阵列。
三、行业影响:从基础研究到产业应用
1. 作物抗病毒育种的新靶点
全球每年因植物病毒导致的农作物损失超过300亿美元。传统抗病毒策略多依赖单一R基因,易被病毒快速突破。DCL4-DRB4通路作为广谱抗病毒机制,提供了全新育种靶点:
增强DCL4表达:通过基因编辑提高作物DCL4活性,增强对多种病毒的抵抗力
优化LinkerRBDs:设计具有更长RNA偏好的DCL4变体,提升对长病毒基因组的处理效率
DRB4协同工程:改造DRB4与DCL4的互作界面,提高复合物稳定性
案例前瞻:中国农科院团队正尝试将水稻DCL4基因导入易感病毒的马铃薯品种,初步实验显示转基因植株对马铃薯Y病毒的抗性提高3.5倍。
2. RNA农药设计的分子指南
基于RNA干扰的农药(如dsRNA喷雾剂)因其高度特异性和环境友好性,被视为下一代绿色农药。然而,成本高、稳定性差是主要瓶颈。本研究为优化植物RNA农药提供了关键指导:
长度优化:设计50-100 bp的dsRNA片段,匹配DCL4-DRB4的底物偏好,提高加工效率
末端修饰:在5'端添加磷酸基团,3'端设计2-nt突出,增强DCL4识别
递送系统:包裹在纳米颗粒中的dsRNA应暴露足够长度的末端,便于DCL4-DRB4捕获
3. 合成生物学工具箱的扩充
DCL4-DRB4的"连续切割"特性,使其成为合成生物学中理想的RNA处理工具:
可编程RNA切割器:融合DCL4切割域与人工RNA识别模块,创建定制化RNA处理器
生物传感器:设计响应特定分子的DCL4变体,切割报告RNA产生可检测信号
基因线路调控:利用DCL4切割特性构建RNA级联放大系统,增强合成基因线路的灵敏度
4. 中国在结构植物学领域的引领地位
该研究由南方科技大学杜嘉木团队主导,与山东农业大学、北京大学现代农业研究院等机构合作完成,标志着中国在植物RNA干扰结构生物学领域已处于国际引领地位。研究使用的技术路线(低温电镜解析RNA-蛋白复合物)为同类研究树立了新标准,其方法论将惠及整个植物分子生物学领域。
四、未来展望
DCL4-DRB4-AGO功能模块解析
下一步需解析DCL4-DRB4与AGO1(siRNA效应器)的复合物结构,揭示siRNA从生成到装载的全过程。
病毒反防御机制的对抗研究
许多植物病毒编码VSRs(病毒抑制RNA沉默蛋白)靶向DCL4,解析这些互作将揭示宿主-病原军备竞赛的分子细节。
作物DCL4的多样性挖掘
水稻、小麦、玉米等主要作物的DCL4可能存在功能分化,系统比较将指导精准作物保护策略。
人工DCL4设计
基于结构模拟,设计具有定制切割长度(如22-nt或24-nt)的DCL4变体,拓展RNA干扰工具箱。
五、结语
在植物细胞的微观世界中,DCL4-DRB4复合物如同一位沉默的守护者,以纳米级的精度"裁剪"RNA,为植物构筑起抵御病毒入侵的分子防线。这项由中国科学家主导的研究,不仅揭示了这一精密分子机器的工作原理,更打开了定向设计植物免疫系统的大门。
从田间地头到实验室,从基础发现到产业应用,RNA干扰技术正悄然改变农业的未来。当科学家们学会"读懂"植物的防御密码,人类与自然的合作将迈入新纪元——不是征服自然,而是与之共舞。这或许是我们从DCL4-DRB4"RNA剪刀匠"身上,获得的最深刻启示。

